KOSIN UNIVERSITY COLLEGE OF MEDICINE
KOSIN UNIVERSITY COLLEGE OF MEDICINE
Articles
- Page Path
- HOME > Kosin Med J > Volume 37(4); 2022 > Article
-
Original article
Correlation of long interspersed element-1 open reading frame 1 and c-Met proto-oncogene protein expression in primary and recurrent colorectal cancers -
Kyung-Yoon Jeon1, Eun-Ji Ko1
 , Hee-Kyung Chang2, Seung-Hyun Lee3, Byung-Kwon Ahn3, Mee Sun Ock1, Hee-Jae Cha1
, Hee-Kyung Chang2, Seung-Hyun Lee3, Byung-Kwon Ahn3, Mee Sun Ock1, Hee-Jae Cha1 -
Kosin Medical Journal 2022;37(4):283-290.
DOI: https://doi.org/10.7180/kmj.22.106
Published online: December 22, 2022
1Department of Parasitology and Genetics, Kosin University College of Medicine, Busan, Korea
2Department of Pathology, Kosin University College of Medicine, Busan, Korea
3Department of Surgery, Kosin University College of Medicine, Busan, Korea
- Corresponding Author: Hee-Jae Cha, PhD Department of Parasitology and Genetics, Kosin University College of Medicine, 262 Gamcheon-ro, Seo-gu, Busan 49267, Korea Tel: +82-51-990-6428 Fax: +82-51-990-3081 E-mail: hcha@kosin.ac.kr
Copyright © 2022 Kosin University College of Medicine.
This is an open-access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
- 1,369 Views
- 26 Download
- 1 Crossref
Abstract
-
Background
- Colorectal cancer is one of the most common cancers worldwide. Colorectal cancer that has recurred and metastasized to other organs also has a very poor prognosis. According to recent studies, the long interspersed element-1 (LINE-1) retrotransposon open reading frame (ORF) is located in the intron of the c-Met proto-oncogene, which is involved in cancer progression and metastasis, and regulates its expression. However, no study has compared the expression patterns of LINE-1 ORF1 and c-Met, which are closely related to cancer progression and metastasis, and their correlation in primary and recurrent cancers.
-
Methods
- In the present study, we compared the expression patterns of LINE-1 ORF1 and c-Met in both primary and recurrent colorectal cancer tissues from 10 patients. Expression patterns and correlations between LINE-1 ORF1 and c-Met proto-oncogene proteins were analyzed by immunofluorescence staining using both LINE-1 ORF1 and c-Met antibodies.
-
Results
- The expression patterns of LINE-1 ORF1 and c-Met showed significant individual differences, and the expression of both proteins was correlated in all colorectal cancer patients. However, the expression levels of LINE-1 ORF1 and c-Met were not significantly different between primary and recurrent colorectal cancers.
-
Conclusions
- The protein expression levels of LINE-1 ORF1 and c-Met were correlated, but did not change significantly in cases of recurrent colorectal cancer in the same patient.
- Colorectal cancer is the third most common cancer in the world, after lung and breast cancer [1-4]. The prevalence rate of people under 50 years is low, but older people tend to have an increased prevalence rate closely related to Westernized eating habits [3,4]. Most colorectal cancers are difficult to completely cure, and recurrences often occur [4]. Metastatic recurrent colorectal cancer usually appears in the liver, lung or in the pelvis and peritoneum with severe symptoms [5].
- Recent studies have shown that transposable elements account for 45% of the total human genome. In particular, long interspersed element-1 (LINE-1 or L1) accounts for 16% to 20% and is associated with the development of cancer [6-8]. LINE-1 consists of a 5′-untranslated region (UTR), a 3′-UTR and two types of open reading frames (ORFs) translated into protein [6,9-11]. ORF1 encodes ~40 kDa nucleic acid binding protein (ORF1p) [12-16] with nucleic acid chaperone activity [16,17]. ORF2 encodes a ~150 kDa protein (ORF2p) with DNA endonuclease [18] and reverse transcriptase activity [19,20].
- LINE-1 is used as a representative measurement of DNA methylation of the entire genome as it belongs to the retrotransposon that exists in the whole human genome [9-11,21,22]. Hypomethylation of LINE-1 has been studied intensively, observed in almost all cancer types, and can be a hallmark of many human cancers [23,24]. LINE-1 hypomethylation induces the activation of proto-oncogenes in human colorectal cancer metastasis. Hypomethylation of LINE-1 is increased in cancer metastasis [7]. Specifically, the activation of proto-oncogene c-Met is involved in liver metastasis of colorectal cancer patients [7]. Moreover, the transcripts of LINE-1 are distributed throughout the genome, and c-Met also contains LINE-1 transcripts [25]. c-Met is over-expressed in colorectal cancer metastases; it was reported that it is regulated by hypomethylation of the internal anti-sense promoter of LINE-1 rather than the promoter site of colorectal cancer metastasis [7,25].
- A well-known oncogene, c-Met proto-oncogene located on chromosome 7 encodes the transmembrane tyrosine kinase, a receptor for hepatocyte growth factor (HGF) or scatter factor (SF) [22,25-33]. When c-Met is activated by binding with a ligand (HGF or SF), it induces various cell reactions such as cell proliferation, motility, scattering, and invasion [28,34-38]. It can also induce invasion through biological effects on epithelial cells [28,30,36]. Based on this knowledge, the c-Met/ligand complex is considered to control the growth and migration of various epithelial cancers [28,29,32,33,35,37,38]. In addition, the c-Met proto-oncogene is over-expressed in cancer and is thought to be an important biomarker [28,32,33,36-38].
- There have been many previous studies on LINE-1 and c-Met. Most were comparisons of protein expression levels and patterns between normal and cancer tissues. These relationships in primary and recurrent cancers derived from the same patient, however, are poorly understood. In this study, we analyzed LINE-1 ORF1 and c-Met proto-oncogene expression levels in both primary and recurrent colorectal cancer tissues from 10 patients.
Introduction
- Ethical statements: This study was approved by the Institutional Review Board of Kosin University Gospel Hospital (IRB No. KUGH IRB 11-60). Each patient involved in the study provided written informed consent.
- 1. Patient sample
- Primary and recurrent cancer paraffin slides from 10 colorectal cancer patients were obtained from the Department of Pathology, Kosin University Gospel Hospital. Personal information such as age, sex, cancer region, and surgery date is presented in Table 1.
- 2. Immunofluorescence analysis
- For immunofluorescence analysis, paraffin slides of 10 patient samples were dried at 60℃ for 1 hour. After that, xylene and ethanol were used to deparaffinize and hydrate the slides. They were then immersed in a citrate buffer (0.01 M, pH 6.0) for antigen retrieval, and heated for 10 minutes in a microwave (700 W or higher) three times. A Dako pen (Dako Denmark, Glostrup, Hovedstaden, Denmark) was used around the cancer tissue to prevent the solution from escaping. To block nonspecific antibodies, slides were treated with 5% fetal bovine serum in 1X phosphate buffered saline (PBS) for 1 hour. After treatment, slides were incubated with anti-LINE-1 ORF1p (1:1,000 dilution; EMD Millipore Corp., Temecula, CA, USA), a mouse monoclonal antibody, and anti-Met (c-Met) antibody (1:500 dilution; Abcam, Cambridge, UK), a rabbit monoclonal antibody, overnight at 4℃. They were then washed three times with 1X PBS for 5 minutes. After that process, slides were incubated with goat anti-mouse secondary antibody Alexa Fluor 546 (1:1,000 dilution; Invitrogen, Carlsbad, CA, USA) and donkey anti-rabbit secondary antibody Alexa Fluor 488 (1:1,000 dilution, Invitrogen) for 1 hour. They were then washed three times with 1X PBS for 5 minutes. For nuclear staining, slides were incubated with Qnuclear Deep Red stain (1:1,000 dilution, Invitrogen) for 20 minutes, then washed four times with 1X PBS for 5 minutes. Specimen epifluorescence was confirmed using a confocal laser scanning microscope (LSM 510 META; ZEISS, Atlanta, GA, USA). The specificity of each antibody was confirmed by staining of mouse polyclonal IgG and rabbit polyclonal IgG. Confocal images were used for measuring fluorescence intensity with image analysis software AlphaEase FC (Alpha Innotech, Randburg, Gauteng, South Africa).
- 3. Statistical analysis
- The statistics were calculated by patient’s experimental results. Fluorescence images were analyzed using AlphaEase FC and image intensity was used for calculation of mean values and 95% confidence intervals. Both values were compared using two-tailed Student t-test. p-values <0.05 were considered statistically significant. Correlation of protein expression was confirmed by SPSS 27 software (IBM Corp., Armonk, NY, USA).
Methods
- 1. Expression patterns of LINE-1 ORF1 and c-Met
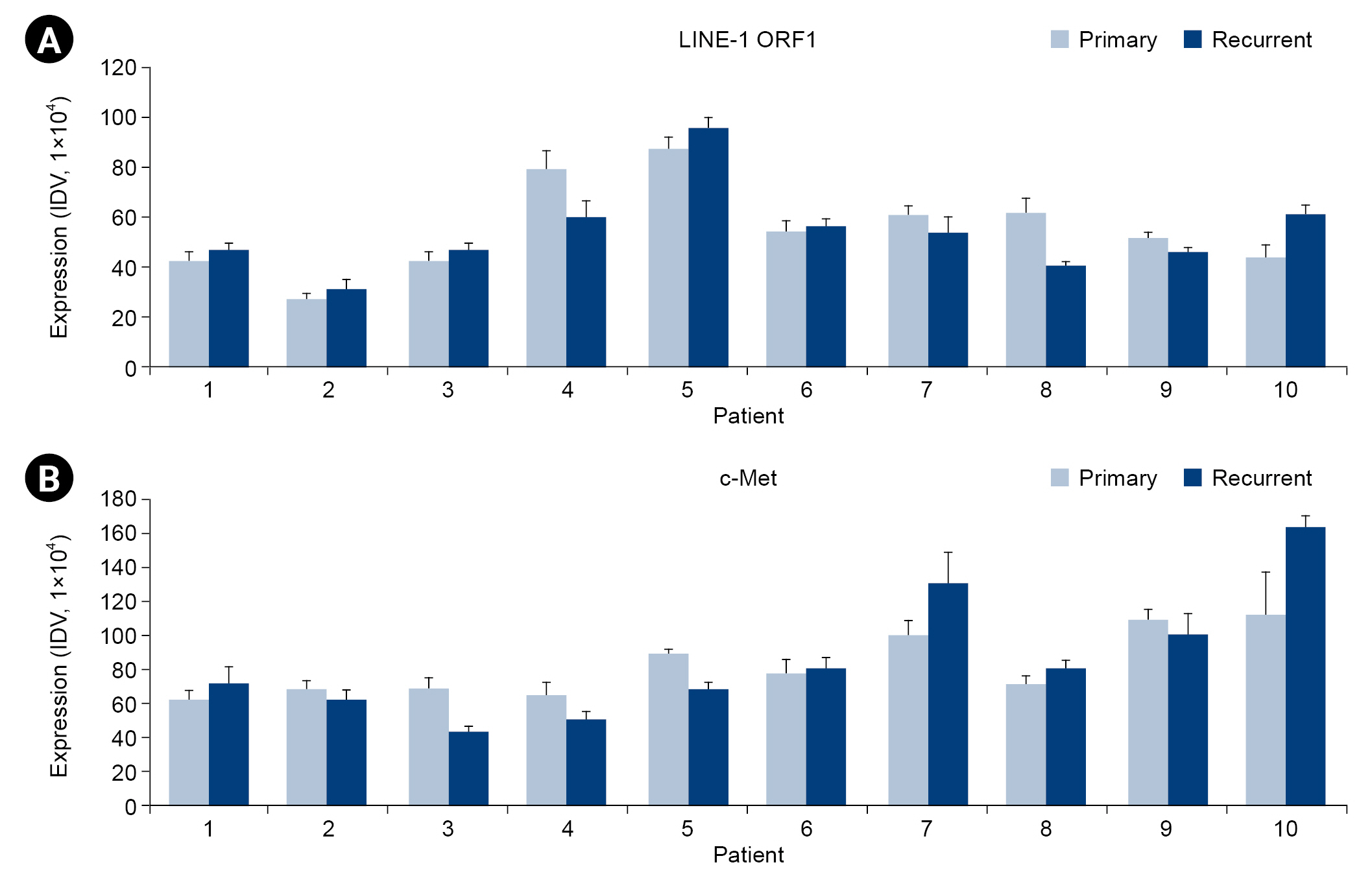
- LINE-1 ORF1 and c-Met proto-oncogene protein expression were confirmed in 10 pairs of patient colorectal cancer samples by immunofluorescence staining (Fig. 1). Each patient’s slide contained primary and recurrent colorectal cancers from the same patient. Immunofluorescence staining image analysis showed that the expression patterns of LINE-1 ORF1 and c-Met varied among patients. Patients 1, 6, 7, 8, and 10 had high expression of c-Met in the recurrent group, and patients 2, 3, 4, 5, and 9 had high expression of c-Met in the primary group. Patients 1, 2, 5, 6, and 10 had high expression of LINE-1 in the recurrent group, and patients 3, 4, 7, 8, and 9 had high expression of LINE-1 in the primary group. These results indicate that the expression pattern is specific to each individual. Expression levels were consistent in both primary and recurrent colorectal cancers from the same patient (Fig. 2).
- 2. LINE-1 ORF1 and c-Met protein expression in primary and recurrent colorectal cancer
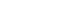
- Based on immunofluorescence staining analysis, the integrated density value was calculated and plotted as a graph (Fig. 3). When the expression of the LINE-1 ORF1 and c-Met proteins in primary and recurrent colorectal cancers was compared in the same patient, overall patterns were similar (Fig. 3).
- 3. Correlation between LINE-1 ORF1 and c-Met expression
- Although the expression levels of LINE-1 ORF1 and c-Met varied among patients, the expression of LINE-1 ORF1 and c-Met were correlated with each other (Fig. 4). On correlation analysis, LINE-1 ORF1 and c-Met protein expression were related to each other in three groups: primary (r=0.203, p<0.05), recurrent (r=0.233, p<0.05), and primary and recurrent (r=0.217, p<0.01) (Table 2).
Results
- LINE-1 is one of the most abundant mobile genetic elements in the human genome and it acts as retrotransposon [9,11,21,22]. Activated LINE-1 causes modification of the original DNA sequence, which negatively affects the stability of the chromosome [6]. Thus, LINE-1 can contribute to the genomic instability that is characteristic of cancer. Moreover, LINE-1 elements are distributed throughout the genome and may affect the expression of marginal genes. A recent study showed that LINE-1 sequences inserted into the c-MET gene intron between exons 2 and 3 and LINE-1 hypomethylation affected c-MET proto-oncogene expression [18,25,39]. A well-known proto-oncogene, c-MET is located on chromosome 7 and is over-expressed in cancer; thus, it has been suggested to be a cancer biomarker [22,25,26,29-33]. LINE-1 and the c-MET proto-oncogene are closely related to cancer and their protein expression levels have been reported to be upregulated in many cases [23,35].
- In this study, LINE-1 ORF1 and c-MET proto-oncogene protein expression levels and expression patterns were analyzed in primary and recurrent colorectal cancer patient samples. We investigated the correlation between the LINE-1 ORF1 and c-Met protein expression in individual patients. Previous reports suggested that LINE-1 ORF1 hypomethylation and c-MET expression are highly related to tumor malignancy and metastatic potential [7]. However, the expression patterns of LINE-1 ORF1 and c-MET protein expression were similar in recurrent cancer compared with primary cancer. The expression pattern of both proteins exhibited individual patterns, meaning that expression varies by patient but is preserved in recurrent cancers in the same patient (Figs. 2, 3). LINE-1 ORF1 and c-Met protein expression were significantly correlated (Table 2, Fig. 4). However, Pearson correlation coefficients were weak. These data suggested that LINE-1 ORF1 and c-Met levels are correlated, but other factors may be involved in regulation of c-Met protein expression. Because this study only examined the whole LINE-1 ORF1 protein in colorectal cancer samples, the expression pattern of specific LINE-1 ORF1 interspersed c-Met genome constructs should be evaluated in the future.
- In conclusion, protein expression levels of LINE-1 ORF1 and c-Met were not significantly different between primary and recurrent colorectal cancers in the same patient. There were interindividual differences in protein expression, while intraindividual patterns were consistent. The expression of both LINE-1 ORF1 and c-Met protein was correlated.
Discussion
-
Conflicts of interest
Hee-Jae Cha is an editorial board member of the journal but was not involved in the peer reviewer selection, evaluation, or decision process of this article. No other potential conflicts of interest relevant to this article were reported.
-
Funding
This work was supported by a National Research Foundation Grant funded by the Korean Government (NRF-2016R1D1A3B01007444).
-
Author contributions
Conceptualization: KYJ. Data curation: EJK. Formal analysis: KYJ. Funding acquisition: HJC. Methodology: KYJ, EJK, HKC, SHL, BKA. Project administration: MSO, HJC. Visualization: EJK, HJC. Writing - original draft: KYJ, HJC. Writing - review & editing: EJK, HKC, SHL, BKA, MSO, HJC. Approval of final manuscript: all authors.
Article information


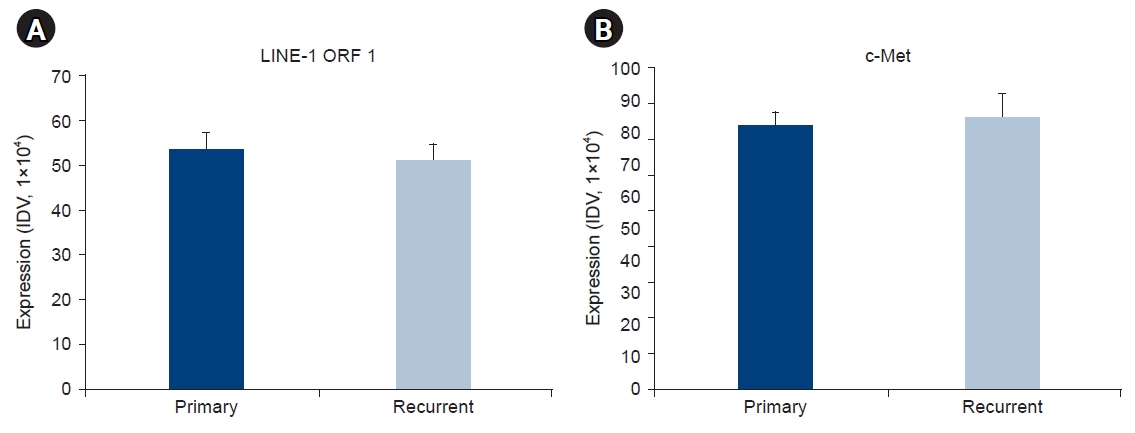

| Correlations | ra) | p-value | No. |
|---|---|---|---|
| Primary | 0.203b) | 0.042 | 101 |
| Recurrence | 0.233b) | 0.018 | 103 |
| Primary & recurrence | 0.217c) | 0.002 | 203 |
- 1. Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer 2010;127:2893–917.ArticlePubMed
- 2. Labianca R, Beretta GD, Kildani B, Milesi L, Merlin F, Mosconi S, et al. Colon cancer. Crit Rev Oncol Hematol 2010;74:106–33.ArticlePubMed
- 3. Brenner H, Kloor M, Pox CP. Colorectal cancer. Lancet 2014;383:1490–502.ArticlePubMed
- 4. Jalili-Nik M, Soltani A, Moussavi S, Ghayour-Mobarhan M, Ferns GA, Hassanian SM, et al. Current status and future prospective of Curcumin as a potential therapeutic agent in the treatment of colorectal cancer. J Cell Physiol 2018;233:6337–45.ArticlePMCPDF
- 5. Kang YJ, Jo JO, Ock MS, Chang HK, Lee SH, Ahn BK, et al. Thymosin β4 was upregulated in recurred colorectal cancers. J Clin Pathol 2014;67:188–90.ArticlePubMed
- 6. Chen L, Dahlstrom JE, Chandra A, Board P, Rangasamy D. Prognostic value of LINE-1 retrotransposon expression and its subcellular localization in breast cancer. Breast Cancer Res Treat 2012;136:129–42.ArticlePubMedPMC
- 7. Hur K, Cejas P, Feliu J, Moreno-Rubio J, Burgos E, Boland CR, et al. Hypomethylation of long interspersed nuclear element-1 (LINE-1) leads to activation of proto-oncogenes in human colorectal cancer metastasis. Gut 2014;63:635–46.ArticlePubMedPMC
- 8. Mima K, Nowak JA, Qian ZR, Cao Y, Song M, Masugi Y, et al. Tumor LINE-1 methylation level and colorectal cancer location in relation to patient survival. Oncotarget 2016;7:55098–109.ArticlePubMedPMC
- 9. Speek M. Antisense promoter of human L1 retrotransposon drives transcription of adjacent cellular genes. Mol Cell Biol 2001;21:1973–85.ArticlePubMedPMCPDF
- 10. Nigumann P, Redik K, Matlik K, Speek M. Many human genes are transcribed from the antisense promoter of L1 retrotransposon. Genomics 2002;79:628–34.ArticlePubMed
- 11. Khalid M, Bojang P Jr, Hassanin AA, Bowers EC, Reyes-Reyes EM, Ramos IN, et al. Line-1: implications in the etiology of cancer, clinical applications, and pharmacologic targets. Mutat Res Rev Mutat Res 2018;778:51–60.ArticlePubMed
- 12. Martin SL. Ribonucleoprotein particles with LINE-1 RNA in mouse embryonal carcinoma cells. Mol Cell Biol 1991;11:4804–7.ArticlePubMedPMC
- 13. Holmes SE, Singer MF, Swergold GD. Studies on p40, the leucine zipper motif-containing protein encoded by the first open reading frame of an active human LINE-1 transposable element. J Biol Chem 1992;267:19765–8.ArticlePubMed
- 14. Hohjoh H, Singer MF. Sequence-specific single-strand RNA binding protein encoded by the human LINE-1 retrotransposon. EMBO J 1997;16:6034–43.ArticlePubMedPMC
- 15. Hohjoh H, Singer MF. Cytoplasmic ribonucleoprotein complexes containing human LINE-1 protein and RNA. EMBO J 1996;15:630–9.ArticlePubMedPMCPDF
- 16. Khazina E, Weichenrieder O. Non-LTR retrotransposons encode noncanonical RRM domains in their first open reading frame. Proc Natl Acad Sci U S A 2009;106:731–6.ArticlePubMedPMC
- 17. Feng Q, Moran JV, Kazazian HH Jr, Boeke JD. Human L1 retrotransposon encodes a conserved endonuclease required for retrotransposition. Cell 1996;87:905–16.ArticlePubMed
- 18. Wolff EM, Byun HM, Han HF, Sharma S, Nichols PW, Siegmund KD, et al. Hypomethylation of a LINE-1 promoter activates an alternate transcript of the MET oncogene in bladders with cancer. PLoS Genet 2010;6:e1000917.ArticlePubMedPMC
- 19. Mathias SL, Scott AF, Kazazian HH Jr, Boeke JD, Gabriel A. Reverse transcriptase encoded by a human transposable element. Science 1991;254:1808–10.ArticlePubMed
- 20. Dombroski BA, Feng Q, Mathias SL, Sassaman DM, Scott AF, Kazazian HH Jr, et al. An in vivo assay for the reverse transcriptase of human retrotransposon L1 in Saccharomyces cerevisiae. Mol Cell Biol 1994;14:4485–92.ArticlePubMedPMCPDF
- 21. Drongitis D, Rainone S, Piscopo M, Viggiano E, Viggiano A, De Luca B, et al. Epigenetics and cortical spreading depression: changes of DNA methylation level at retrotransposon sequences. Mol Biol Rep 2016;43:755–60.ArticlePubMedPDF
- 22. Burns KH. Transposable elements in cancer. Nat Rev Cancer 2017;17:415–24.ArticlePubMedPDF
- 23. Rodic N, Sharma R, Sharma R, Zampella J, Dai L, Taylor MS, et al. Long interspersed element-1 protein expression is a hallmark of many human cancers. Am J Pathol 2014;184:1280–6.ArticlePubMedPMC
- 24. Swets M, Zaalberg A, Boot A, van Wezel T, Frouws MA, Bastiaannet E, et al. Tumor LINE-1 methylation level in association with survival of patients with stage II colon cancer. Int J Mol Sci 2016;18:36.ArticlePubMedPMC
- 25. Miglio U, Berrino E, Panero M, Ferrero G, Coscujuela Tarrero L, Miano V, et al. The expression of LINE1-MET chimeric transcript identifies a subgroup of aggressive breast cancers. Int J Cancer 2018;143:2838–48.ArticlePubMedPDF
- 26. Park M, Dean M, Kaul K, Braun MJ, Gonda MA, Vande Woude G. Sequence of MET protooncogene cDNA has features characteristic of the tyrosine kinase family of growth-factor receptors. Proc Natl Acad Sci U S A 1987;84:6379–83.ArticlePubMedPMC
- 27. Birchmeier C, Birchmeier W, Gherardi E, Vande Woude GF. Met, metastasis, motility and more. Nat Rev Mol Cell Biol 2003;4:915–25.ArticlePubMedPDF
- 28. Ma PC, Maulik G, Christensen J, Salgia R. c-Met: structure, functions and potential for therapeutic inhibition. Cancer Metastasis Rev 2003;22:309–25.PubMed
- 29. Kammula US, Kuntz EJ, Francone TD, Zeng Z, Shia J, Landmann RG, et al. Molecular co-expression of the c-Met oncogene and hepatocyte growth factor in primary colon cancer predicts tumor stage and clinical outcome. Cancer Lett 2007;248:219–28.ArticlePubMed
- 30. Boccaccio C, Comoglio PM. MET, a driver of invasive growth and cancer clonal evolution under therapeutic pressure. Curr Opin Cell Biol 2014;31:98–105.ArticlePubMed
- 31. Chen T, Cai SL, Li J, Qi ZP, Li XQ, Ye LC, et al. Mecp2-mediated epigenetic silencing of miR-137 contributes to colorectal adenoma-carcinoma sequence and tumor progression via relieving the suppression of c-Met. Sci Rep 2017;7:44543.ArticlePubMedPMCPDF
- 32. Scheri KC, Leonetti E, Laino L, Gigantino V, Gesualdi L, Grammatico P, et al. c-MET receptor as potential biomarker and target molecule for malignant testicular germ cell tumors. Oncotarget 2018;9:31842–60.ArticlePubMedPMC
- 33. Zhang J, Babic A. Regulation of the MET oncogene: molecular mechanisms. Carcinogenesis 2016;37:345–55.ArticlePubMed
- 34. Di Renzo MF, Olivero M, Giacomini A, Porte H, Chastre E, Mirossay L, et al. Overexpression and amplification of the met/HGF receptor gene during the progression of colorectal cancer. Clin Cancer Res 1995;1:147–54.PubMed
- 35. Ma PC, Tretiakova MS, MacKinnon AC, Ramnath N, Johnson C, Dietrich S, et al. Expression and mutational analysis of MET in human solid cancers. Genes Chromosomes Cancer 2008;47:1025–37.ArticlePubMedPMC
- 36. Zeng ZS, Weiser MR, Kuntz E, Chen CT, Khan SA, Forslund A, et al. c-Met gene amplification is associated with advanced stage colorectal cancer and liver metastases. Cancer Lett 2008;265:258–69.ArticlePubMedPMC
- 37. Boromand N, Hasanzadeh M, ShahidSales S, Farazestanian M, Gharib M, Fiuji H, et al. Clinical and prognostic value of the C-Met/HGF signaling pathway in cervical cancer. J Cell Physiol 2018;233:4490–6.ArticlePubMedPDF
- 38. Luo Y, Ouyang J, Zhou D, Zhong S, Wen M, Ou W, et al. Long noncoding RNA GAPLINC promotes cells migration and invasion in colorectal cancer cell by regulating miR-34a/c-MET signal pathway. Dig Dis Sci 2018;63:890–9.ArticlePubMedPDF
- 39. Zhu C, Utsunomiya T, Ikemoto T, Yamada S, Morine Y, Imura S, et al. Hypomethylation of long interspersed nuclear element-1 (LINE-1) is associated with poor prognosis via activation of c-MET in hepatocellular carcinoma. Ann Surg Oncol 2014;21 Suppl 4:S729–35.ArticlePubMedPDF
References
Figure & Data
References
Citations
- Functional Analysis of Membrane-Associated Scaffolding Tight Junction (TJ) Proteins in Tumorigenic Characteristics of B16-F10 Mouse Melanoma Cells
Eun-Ji Ko, Do-Ye Kim, Min-Hye Kim, Hyojin An, Jeongtae Kim, Jee-Yeong Jeong, Kyoung Seob Song, Hee-Jae Cha
International Journal of Molecular Sciences.2024; 25(2): 833. CrossRef
 PubReader
PubReader ePub Link
ePub Link Cite
Cite